Effects of aerobic exercise on the expressions of skeletal muscle fiber type-related genes and FNDC-5 in high-fat diet-induced obese rats
International Journal of Applied Sports Sciences, Vol.29, No.1, pp.42-53
https://doi.org/10.24985/ijass.2017.29.1.42
ⓒ Korea Institute of Sport Science
초록
Obesity is known to be linked with a shift in skeletal muscle fiber type, which plays a role in insulin sensitivity and myokines activity. Here, we investigated whether aerobic exercise training has any effects on skeletal muscle MHC isoforms related to gene expression, irisin, and FNDC5 in obese rats. Thirty-two male SD-rats (3 wks old) were randomly assigned to normal diet (ND, n = 8) and high-fat diet (HFD, n = 24) groups, with specific caloric intake (ND; 11.5%, HFD; 60% of kcals from fat), for 8 weeks. After 8 weeks, HFD-induced obese rats were randomly reassigned to either an obese control (OC, n = 8) or obese exercise (OE, n = 8) group, and provided a normal diet. The OE group was exercised on the rodent treadmill (60 min, 15 m/min, 5 times) for 8 weeks. We investigated FNDC5, slow-twitch muscle gene (miR-499) and fast-twitch muscle gene (SOX6) expression in gastrocnemius muscle, and serum irisin. HFD group showed heavier body weight compared to ND group (p < .05). Blood glucose concentrations, determined by IGTT, were significantly higher in HFD group compared to ND group in 90-minutes and 120-minutes of the recovery phase (p < .05, 43%, 34%). Average body weight for the OE group was significantly lower than the OC group. The level of blood glucose in the OE group was lower than the OC group in 90-minutes and 120-minutes of the recovery phase (p < .05, 11%, 9%). Irisin levels were not significantly different between the HFD and ND groups. However, FNDC5 level was significantly lower in HFD rats than ND (p < .05, 53%). The OE group showed increased expression of irisin and FNDC5 compared to ND, but there was no change in OC (p < .05, 23%, 42%). While the levels of miR-499 and SOX6 were significantly higher in the OE compared to OC group, there was no change in MYH7. Taken together, 8 week regular aerobic exercise has positive effects on body weight, blood irisin concentration, its precursor FNDC5, as well as fiber type promoting genes mi-499 and SOX6.
Abstract
Obesity is known to be linked with a shift in skeletal muscle fiber type, which plays a role in insulin sensitivity and myokines activity. Here, we investigated whether aerobic exercise training has any effects on skeletal muscle MHC isoforms related to gene expression, irisin, and FNDC5 in obese rats. Thirty-two male SD-rats (3 wks old) were randomly assigned to normal diet (ND, n = 8) and high-fat diet (HFD, n = 24) groups, with specific caloric intake (ND; 11.5%, HFD; 60% of kcals from fat), for 8 weeks. After 8 weeks, HFD-induced obese rats were randomly reassigned to either an obese control (OC, n = 8) or obese exercise (OE, n = 8) group, and provided a normal diet. The OE group was exercised on the rodent treadmill (60 min, 15 m/min, 5 times) for 8 weeks. We investigated FNDC5, slow-twitch muscle gene (miR-499) and fast-twitch muscle gene (SOX6) expression in gastrocnemius muscle, and serum irisin. HFD group showed heavier body weight compared to ND group (p < .05). Blood glucose concentrations, determined by IGTT, were significantly higher in HFD group compared to ND group in 90-minutes and 120-minutes of the recovery phase (p < .05, 43%, 34%). Average body weight for the OE group was significantly lower than the OC group. The level of blood glucose in the OE group was lower than the OC group in 90-minutes and 120-minutes of the recovery phase (p < .05, 11%, 9%). Irisin levels were not significantly different between the HFD and ND groups. However, FNDC5 level was significantly lower in HFD rats than ND (p < .05, 53%). The OE group showed increased expression of irisin and FNDC5 compared to ND, but there was no change in OC (p < .05, 23%, 42%). While the levels of miR-499 and SOX6 were significantly higher in the OE compared to OC group, there was no change in MYH7. Taken together, 8 week regular aerobic exercise has positive effects on body weight, blood irisin concentration, its precursor FNDC5, as well as fiber type promoting genes mi-499 and SOX6.
Introduction
Human skeletal muscle fibers are composed of type I, type IIa, type IIx fibers, distinguished by Myosin Heavy Chain (MHC) isoforms (Schiaffino & Reggiani, 2011; Liu et al, 2015). However, it is practically divided into 5 fiber types; type I, I/IIa, IIa, IIa/IIx, and I/IIa/IIx. Rodents have a total of 12 types of MHC isoforms including type I, I/IIa, IIa, IIa/IIx, IIx, IIx/IIb, IIb,. MHC isoforms plays a central role in muscle plasticity, such as maximal contraction force and velocity, fatigue resistance, and calcium sensitivity (Widrick et al; 1998; Widrick, 2002). The characteristics of type I muscle fiber is the higher number of mitochondria, and it relies mainly on mitochondrial oxidative metabolism for ATP production (Schiaffino & Reggiani, 2011; Liu et al, 2015). On the contrary, type II muscle fiber has lower mitochondria and is involved in glycolytic metabolism (Schiaffino & Reggiani, 2011; Liu et al, 2015).
Obesity is closely related to metabolic diseases such as high blood pressure, hyperlipidemia, and hyperglycemia. Obesity also induces not only an increase in insulin resistance, but also decrease of muscular strength and mass due to the increase of inflammatory factors and change of molecular mechanism (de Wilde et al., 2008). Recently, the correlation between obesity and fiber type was reported (Stuart et al., 2013), showing similarities with disuse muscle atrophy, where type I muscle fiber decreases selectively. As a result, distribution of type II muscle fiber MHC isoforms tend to be more prevalent in obese skeletal muscles; while this decrease of type I muscle fibers results in diminished glucose uptake, insulin sensitivity, and energy production (ATP production) (Stuart et al., 2013; Marin et al., 1994). In other words, there is an inverse correlation between the level of type I muscle fibers and body fat mass (Lithell et al., 1981; Oberbach et al., 2006; Stuart et al., 2013).
Recently, it has been suggested that exercise may have an effect on regulating muscle fiber type resulting from obesity (Kurdiova et al., 2014). It is also well established that aerobic exercise promotes mitochondrial oxidative capacity and increases type I muscle fiber, which elicits higher aerobic capacity. However, some studies reported that muscular oxidative capacity was altered with no shift of muscle fiber types in diet-induced obese mice (Shortreed et al. 2009; Trajcevski et al. 2013). Other studies showed increased slow-twitch muscle fiber with decreased muscle mass in leptin deficient ob/ob mice (Warmington et al., 2000; Kemp et al., 2009). Thus, fiber type related gene expression needs to be investigated to clarify the relationship between aerobic exercise and fiber type shift. Micro RNA-499 (miR-499), an important factor for fiber type shifting, increases the type I MHC isoform by down-regulating transcription factor which inhibits gene expression of fast-twitch muscle fiber contractile protein (Bell et al., 2010; Quiat et al., 2011). In addition, miR-499 is positively related to oxidative capacity such as maximal phosphorylation capacity, and VO2max (Gan et al., 2013).
Meanwhile, Boström et al discovered the hormone irisin in the human body in 2012. Irisin is composed of 122 amino acids, and released in the blood by proteolytic cleavage of the membrane protein, FNDC5 (fibronectin type III domain containing 5) (Jeremic et al., 2017). These factors are controlled by the PGC-1α (peroxisome proliferator-activated receptor Υ coactivator-1 α) protein (Norheim et al., 2014). Aerobic exercise increases expression of PGC-1α in muscle, which induces proteolytic cleavage of FNDC5 and release of irisin. Subsequently, the increased irisin level induces browning of white adipose tissue through expression of UCP1 protein inside mitochondria (Boström et al., 2012; Zafrir, 2013; Kurdiova et al., 2014). Browning is reported to play a role in preventing metabolic disease-related obesity and reducing insulin resistance by increasing energy consumption. Therefore, fat browning is regarded as an effective medium for improving obesity (Huh et al., 2012; Stengel et al., 2013; Roca-Rivada et al., 2013). However, the blood irisin concentration was found to be increased in obesity (Stengel et al., 2013; Crujeiras et al., 2014). On the contrary, Moreno-Navarrete et al. (2013) reported a negative relationship between irisin concentration and obesity. Moreover, there are few studies that observed irisin, and FNDC5 level with fiber type shift in skeletal muscle following regular aerobic exercise on obese subjects. In particular, there is no study, to our knowledge, that investigates the gene expression difference between slow and fast MHC isoforms in skeletal muscles. Therefore, the purpose of this study was to examine the effect of 8 weeks regular aerobic exercise on the fiber type related gene expression, fat browning factors, and exercise capacity in high-fat diet-induced obese rats.
Methods
Animals and experimental design
Male Sprague-Dawley rats (n=32, 3 weeks) were randomly assigned to a ND (normal diet, n=8) and HFD group (high-fat diet, n=24) after a one week acclimatization period. Apart from diet, all animals were housed under the same environmental conditions for 8 weeks. Total calorie consumption for the ND group consisted of 67.7% carbohydrate, 11.5% fat, and 20.8% protein (D10001, Research diet). The HFD group consumed 20% protein, 60% fat, and 20% carbohydrate (D12492, Research diet). Following the 8 weeks diet treatment, 8 rats in each group were sacrificed. The remaining animals from the HFD group were randomly re-assigned to an obese control (OC, n=8) group, and obese exercise (OE, n=8) group for an 8 weeks aerobic exercise treatment. After the second intervention, the 8 rats from each group (OC and OE) were sacrificed (Figure 1). Temperature of the breeding room was 21℃, and lighting was controlled to create a 12-h dark cycle and 12-h light cycle. All animal care and use procedures were approved by the Institutional Animal Care and Use Committee of Inje Medical School.
Figure 1.
Experimental procedure. ND: normal diet, HFD: high-fat diet, OC: obese control, OE: obese exercise
Exercise protocol and tissue preparation
The obese exercise group animals performed the aerobic exercise at 15 m/min on a 0 % grade for 60 min, 5 days a week for 8 weeks. A Total of 16 animals from the ND (n=8) and HFD (n=8) groups were sacrificed after the 8-week diet treatment, and the remaining 16 rats were sacrificed after 8 weeks of aerobic exercise treatment. All rats were anesthetized by intraperitoneal injection pentobarbital (5mg/100g of body mass). The intact gastrocnemius muscle from the animals were obtained and quick-frozen to be stored at –80℃ until the analysis.
Body Weight measurement
Body weight was measured once a week at a designated time, using an animal scale (A&D Company Limited CE, Japan). Also, 400g of food was supplied once a week to measure the weekly intake.
Glucose tolerance test
2g/kg of glucose was administered by intraperitoneal injection after 12-hour fasting. The blood glucose levels were obtained and measured via tail bleeding, using Accu-check Aviva glucose tips in 30, 60, 90, and 120 minutes after the injection.
Plasma measurements
Blood samples were obtained after 12 hours of overnight fasting using a 10 ml disposable syringe for the blood irisin level analysis. Collected blood was transferred by syringe into EDTA tube, and centrifuged for 10 min at 3000 rpm. After the centrifuging, the supernatant was aliquoted to 1.5ml microtubes and immediately moved to a blood research institute (Seegen medical, Seoul, Korea) for irisin analysis. Plasma irisin (catalog # EK-067-29) was measured using commercial enzyme linked immune sorbent assays kit (ELISA, Phonix Pharamaceuticals, Inc. Burlingame, CA, USA).
RNA extraction
For the extraction of total RNA, Trizol was cultivated for 5 minutes and put to rest at room temperature for three minutes after adding 200μl of chloroform (Sigma, USA). Through centrifugation (13,000rpm, 4℃, 15 minutes), supernatant was separated into a new tube, and the same amount of isopropanol (Sigma, USA) was added. After cultivation at room temperature for 10 minutes, centrifugation (13,000rpm, 4℃, 10 minutes) was conducted. 1 ml of 70% ethanol was added to an RNA pellet that was formed in the bottom of the tube through centrifugation, and the pellet was washed twice (4,500rpm, 4℃, 5 minutes). Once the RNA pellet becomes completely dry, 150 μl of distilled water under 0.01% DEPC treatment was added to dissolve the RNA pellet.
RT-PCR (Reverse transcription-polymerase chain reaction)
For the measurement of mRNA, approximately 14μl of complementary DNA was finally prepared after combining 2μg of total RNA with 0.5μg of reverse primer, and cDNA went under thermal treatment at 75℃ for 15 minutes. The complementary DNA was stored in ice for five minutes. Then, 5X M-MLV reaction buffer, 10mM dNTP mixture, 200 units M-MLV RT (Promega, USA) was added in cDNA to make the final amount 25μl and went under reaction at 42℃ for an hour. 10X reaction buffer (including Mg2+), 10mM dNTP, 5 unites Taq polymerase (Solgent, Korea), and 10μM primer were added to 5μl of RT reaction to make the final amount 50μl. Quantitative Real-time PCR was detected using Tagman Universal Master Mix and 7900HT Sequence Detection System according to the manufacturer’s instructions (Applied Biosystems, Foster City, CA) and detection, and calculation of the cycle threshold (Ct) compared with GAPDH. Primers for miR-499 (Forward: 5′-CTCAACTGGTGTCGTGGAGT-3′, Reverse: 5′-ACACT CCAGCTGGGTTAAGACTTGC-3′), SOX6 (Forward: 5'-GCGCTCACGAAGGCAG-3', Reverse: 5'-GCCATAGTA ATAGCACCAGGATACA-3'), GAPDH (Forward: 5′-GGCT CTCTGCTCCTCCCTGTT-3′, Reverse: 5′-GCGGGATCTCG CTCCTGGAAG -3′) were designed and purchased from Bioneer (Daejon, KOREA).
Western blot
Intact whole gastrocnemius muscle was homogenized by sonication on ice in RIPA buffer (Sigma Aldrich, St. Louis, MO] including protease/phosphatase inhibitor cocktail (Thermo Scientific, Rockford, IL). The homogenate was placed for 30 minutes at 4℃ and centrifuged at 14,000 rpm for 15 min at 4℃, and the supernatant was obtained for analysis. Protein concentration was evaluated using BCA assay Kit (Bio-rad, USA). After 30μg of protein denaturation in loading buffer, samples were loaded onto 9% SDS-PAGE gels and blotted to PVDF membrane. All blots were determined equal loading volume with Ponceau S (Sigma, St Louis, MO) and washed 3 times for 30 min. As for the nonspecific reaction of membrane, the blots were incubated in Tris-buffered saline plus Tween-20 with 5% non-fat dry milk at room temperature for an hour. Primary antibody (ab131390, Abcam, USA) was rabbit polyclonal FNDC-5 at 1:1000 and dissolved in Tris-buffered saline plus Tween-20 with 5% non-fat dry milk at 4℃ for 18 h. The next day, secondary antibody Rabbit-anti-rat IgG (1:1000; Cell Signaling Technology, Danvers, MA) was used for 1 h at room temperature, and the blots were incubated for 5 min with ECL substrate (Pierece Biotechnology). The bands were detected using Kodak film and quantified with Image J software.
Results
Weight change by high-fat diet and aerobic exercise treatment
Baseline weight was not significantly different etween the ND and HFD groups (296.0±3.7g and 293.5±2.0g, respectively). From the 4th week, the HFD group showed an increased body weight compared to the ND group. A significant between-groups difference was observed in the 8th week following the diet treatment where the weight of the ND group was 579.1±19.3g and HFD group was 633.4±13.8g (p<0.05).
Exercise ability following 8 week aerobic exercise treatment
Before the aerobic exercise treatment, the running time up to exhaustion was about 11.2±0.6 minutes both in OC and OE groups. After the 8-week aerobic exercise treatment, the OE group (21.1±2.6 min) had a longer running time than the OC group (15.8±1.1) (p<0.05). Running distance before the experiment treatment was 139±11.5 meters in the OC group and 158±17.1 meters in the OE group, which was not significantly different between groups. After the 8-week aerobic exercise treatment, the OE group (400.3±77.0 meter) recorded a longer exercise distance than the OC group (242.3±22.3 meter) (p<0.05).
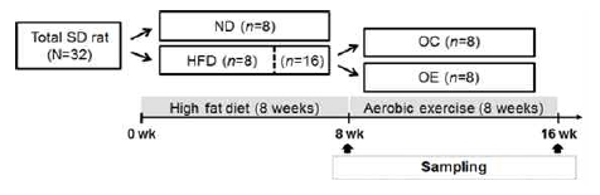
Glucose tolerance following the high-fat diet and aerobic exercise treatment
Blood glucose concentration was 64% higher in the HFD group (14.59 mM) than ND group (8.88 mM) at the recovery period (90 min), as well as being 56% higher in the HFD group (12.43 mM) than the ND group (7.98 mM) after IGTT (p<.05).
HFD-induced obese rats were randomly assigned to an OC group (n=8) and OE group (n=9). The OE group performed aerobic training for 8 weeks. In IGTT test results, OE group blood glucose level (5.64 mM) was 12% lower than that of the OC group (6.38 mM) at 90-minute of the recovery phase after the glucose administration. Likewise, blood glucose level of the OE group (5.35 mM) was 10% lower than the OC group (5.91mM) at 120-minute of the recovery phase (p<.05).
Table 1.
Changes of weight induced by high-fat diet
Table 2.
Changes of weight during aerobic exercise training
Table 3.
Changes of exhausted time and distance by aerobic exercise training
Figure 2.
ND: normal diet (n=8), HFD: high-fat diet (n=9), OC: obese control (n=8), OE: obese exercise (n=9). †indicate a significantly difference between groups.
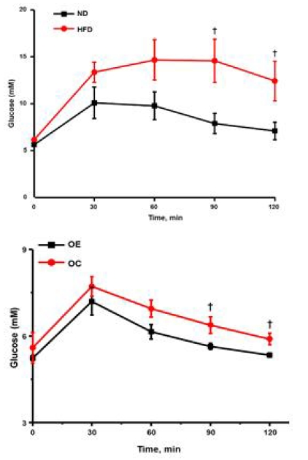
Irisin concentration following the high-fat diet and aerobic exercise
Blood irisin concentration of the HFD group was 739.19±59.22 ng/mL, which is approximately 13.0% lower than the ND group (853.46±99.72 ng/mL). However, there was no statistically significant difference (p>.05). Blood irisin concentration of the OE group, that performed 8-week regular aerobic exercise after inducing obesity, was 957.80±57.18 ng/mL, which is approximately 9% higher than the OC group (879.01±83.87 ng/mL) (p<.05).
Figure 3.
ND, normal diet; HFD, high-fat diet; OC, obese control; OE, obese exercise, Values are Mean ± SE, †indicate a significantly difference between groups
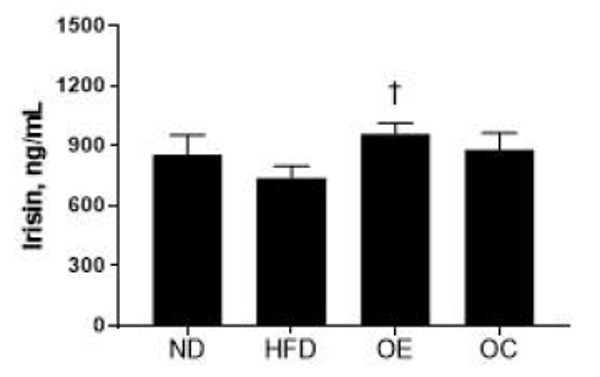
Figure 4.
ND, normal diet; HFD, high-fat diet; OC, obese control; OE, obese exercise, Values are Mean ±SE. †indicate a significantly difference between groups.
FNDC5 expression following the high-fat diet and aerobic exercise treatment
Expression of FNDC5 in the HF group was relatively 55% lower than the ND group (p<0.05). FNDC5 expression of the OE group was 60% higher than the OC group (p<0.05).
Genetic factors following the high-fat diet and aerobic exercise treatment
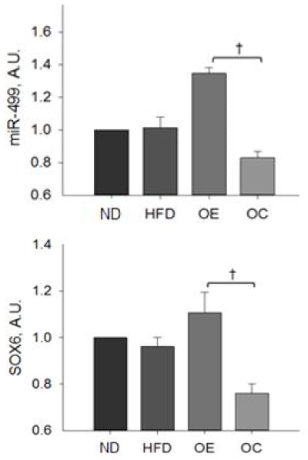
MiR-499 expression of the HFD group was not significantly different compared to the ND group. 8 weeks of regular aerobic exercise, after inducing obesity, resulted in a 64% higher miR-499 expression in the OE group compared to OC group.
Expression of SOX6 protein was lower in the HFD group than in ND group, but there was no significant difference (p<.05). On the other hand, the OE group, after exercise training, showed 43% higher levels of SOX6 expression than the OC group (p<0.05).
Figure 5.
ND, normal diet; HFD, high-fat diet; OC, obese control; OE, obese exercise, Values are Mean ±SE. †indicate a significantly difference between groups.
Discussion
In the current study obesity was induced by an 8-week high-fat diet, and followed by another 8-week aerobic exercise intervention to examine the effects of high-fat diet and aerobic exercise on the shift of muscle fiber isoforms and fat browning. Slow muscle gene, miR-499, fast muscle gene, SOX6, and the proteins that affect fat browning, including irisin and FNDC5 were examined. The following summarizes the major results of this study. First, 8 weeks of high-fat diet resulted in weight gain and reduction of FNDC5, the irisin precursor protein. However, no difference of muscle fiber type was observed. Second, performing 8 weeks of aerobic exercise on obese rats reduced weight. Meanwhile, the exercise induced higher expression of irisin, FNDC5, and also increased miR-499, and SOX6 expression.
It is well known that excessive food intake causes obesity to humans and animals, particularly, greater fat intake results in increased weight gain (Buettner et al., 2007, Ghibaudi et al., 2002). Obesity is also caused by unbalanced energy consumption, and it can cause other metabolic diseases. Hence, it has attracted global attention as a serious health risk. In this study, a high-fat diet (protein 20%, fat 60%, and carbohydrate 20%) was fed in a self-service manner for 8 weeks to SD rats. The results demonstrated a greater weight gain in the HFD group than the ND group (p<.05). In the injection glucose tolerance test, blood glucose concentration of the HFD group was higher than the ND group (p<.05) in 90-minutes and 120-minutes of the recovery phase (64% and 56%, respectively); indicating higher insulin resistance in the HFD group. Weight gain due to a high-fat diet is known to be related to a decrease in metabolic rate (Galgani et al., 2008), fat oxidation (Houmard et al., 2008), decreased activation of the sympathetic nervous system, and increased fat storage (Horton et al., 1995). Whereas, 8 weeks of aerobic exercise treatment induced weight loss significantly greater in the OE group than OC group (p<.05). Moreover, injection glucose tolerance test values were also lower in the OE group than OC group (p<.05) in 90-minutes and 120-minutes of the recovery phase (12% and 10%, respectively). These results imply that regular aerobic exercise can improve insulin sensitivity. Skeletal muscles are a metabolically important organ, as they processes up to 70-90% of glucose after a meal, and are able to process glucose more effectively as a result of exercise (Yu et al., 2016; Nishizawa et al., 2004).
Recently, irisin has gained attention as a potential indicator for improving insulin sensitivity of skeletal muscles, and preventing metabolic disease-related obesity. Irisin is a type of myokine that is secreted in skeletal muscles, and it is known to be secreted depending on the PGC-1a transcription expression (Boström et al. 2012). It has been reported that fat browning due to irisin secretion increases fatty acid in the mitochondria, and glucose oxidation rate (Bartelt et al., 2011). Also it increases energy consumption and AMPK activation, as well as enhancing the efficiency of metabolic factors (Kraemer et al., 2014; Timmons et al., 2012); which can decrease body fat mass (Stengel et al., 2013). In this study, the HFD group showed no difference in blood irisin concentration compared to the ND group. However, the precursor expression of FNDC5 was significantly lower than in the ND (p<.05; 55%). In previous studies, both patients with type II diabetes (Choi et al., 2013) and obesity (Moreno-Navarrete et al. 2013) showed lower blood irisin levels than healthy people. On the contrary, several previous studies observed higher blood irisin concentration in obese patients than healthy people (Stengel et al., 2013; Crujeiras et al., 2014). Therefore, it is hard to draw a clear conclusion on the role of irisin in obesity. In this study, 8 weeks of aerobic exercise treatment to obesity-induced rats (OE group) significantly increased irisin and FNDC5 expression compared to the OC group (p<.05; 9% and 60%, respectively). These results were consistent with a previous study which reported that 10 weeks regular aerobic exercise, on adults, significantly increased blood irisin levels (Boström et al., 2012). It also agrees with reports that aerobic exercise or high-intensity sprints increased body irisin levels (Timmons et al., 2012; Kraemer et al., 2014). Taken together, blood irisin level can be increased by regular aerobic exercise. Lee et al. (2014) also reported an increase of blood irisin concentration after a 60-minute bicycle ride at 40% VO2 max. However, the study reported maintained blood irisin concentration after the maximum exercise test, arguing that irisin concentration is affected by the exercise intensity rather than exercise duration.
Insulin sensitivity of skeletal muscles is stronger with the increase of type I fiber ratio. In other words, the insulin sensitivity is affected by the distribution of muscle fiber type, and it controls synthesis of glycogen in the muscle (Hickey et al., 1995; Tanner et al., 2002). Type II diabetes patients experience selective loss of type I fibers (slow muscle), and it subsequently impairs glucose uptake, insulin sensitivity, and energy production (ATP production) processes (Stuart et al., 2013; Marin et al., 1994). According to Hyatt et al. (2016), high-fat/high-sugar diet treatment for two years on 24-month-old rhesus monkeys increased the mRNA, and altered the muscle fiber type from slow-twitch to fast-twitch in soleus muscle. However, the high-fat/ high-sugar diet group showed no alteration of miR-499, the gene responsible for stimulating slow-twitch muscle fiber expression, or SOX6, the gene stimulating fast-twitch muscle expression. Even though the current study showed no significant changes in miR-499 and SOX6 expression following the aerobic exercise, it was significantly different between groups. In particular, miR-499 (p<0.05, 64%) and SOX6 (p<0.05, 43%) expression in the OE group was higher than the OC group in obesity-induced rats. These results are in alignment with reports that combined treatment of diet restriction and physical activity decreased weight and increased type I fiber composition in quadriceps femoris of obese female (Gerrits et al., 2010). However, it conflicted with a previous study showing a difference in muscle mass with no change of muscle fiber types after 6 weeks of running exercise in mouse skeletal muscle (Glaser et al., 2010). Recently, DeNies et al. (2014) reported that slow-twitch fibers of soleus muscle were reduced with no change of muscle mass in C57BLK/6J mice after inducing obesity by a high-fat diet; which showed an inverse correlation between type I fiber and fat mass. In addition, it was speculated that reduced type I muscle fibers may be connected to increments in type I/IIa hybrid fibers, which imply that type I muscle fibers may easily be transformed to hybrid muscle fibers in male obese mice. These inconsistencies could be attributed to the different diet composition, treatment period, and different species, thus additional studies are needed to clarify the relationship between aerobic exercise and fiber type shift.
In conclusion, 8 weeks of aerobic exercise after a high-fat diet resulted in weight loss, and improved exercise capacity. Moreover, regular aerobic exercise has a positive impact on blood irisin concentration, and expression of FNDC5. The level of miR-499 and SOX6 expression, which are the promoting genes of slow-twitch muscle and fast-twitch muscle, respectively, tend to be increased after the aerobic exercise.
* This work was done with the sports promotion fund from Korea Institute of Sport Science, Korea Sports Promotion Foundation (KISS-15-A03003)
References
Bartelt. A., Bruns, O. T., Reimer, R., Hohenberg, H., Ittrich, H., Peldschus, K., Kaul, M. G., Tromsdorf, U. I., Weller, H., Waurisch, C., Eychmüller, A., Gordts, P. L., Rinninger, F., Bruegelmann, K., Freund, B., Nielsen, P., Merkel, M., & Heeren, J. (2011). Brown adipose tissue activity controls triglyceride clearance. Nature Medicine, 17(2), 200-5.
Boström, P., Wu, J., Jedrychowski, M. P., Korde, A., Ye, L., Lo, J. C., Rasbach, K. A., Boström, E. A., Choi, J. H., Long, J. Z., Kajimura, S., Zingaretti, M. C., Vind, B. F., Tu, H., Cinti, S., Højlund, K., Gygi, S. P., & Spiegelman, B. M. (2012). A PGC1-α-dependent myokine that drives brownfat- like development of white fat and thermogenesis. Nature, 481(7382), 463-8.
Choi, S. J., Shively, C. A., Register, T. C., Feng, X., Stehle, J., High, K., Ip, E., Kritchevsky, S. B., Nicklas, B., & Delbono, O. Force-generation capacity of single vastus lateralis muscle fibers and physical function decline with age in African green vervet monkeys. (2013). Journals of Gerontology. Series A: Biological Sciences and Medical Sciences, 68(3), 258-67.
Crujeiras, A. B., Pardo, M., Arturo, R. R., Navas-Carretero, S., Zulet, M. A., Martínez, J. A., & Casanueva, F. F. (2014). Longitudinal variation of circulating irisin after an energy restriction-induced weight loss and following weight regain in obese men and women. American Journal of Human Biology, 26(2), 198-207.
Gerrits, M. F., Ghosh, S., Kavaslar, N., Hill, B., Tour, A., Seifert, E. L., Beauchamp, B., Gorman, S., Stuart, J., Dent, R., McPherson, R., Harper, M.E. (2010). Distinct skeletal muscle fiber characteristics and gene expression in diet-sensitive versus diet-resistant obesity. The Journal of Lipid Research, 51(8), 2394-404.
Huh, J. Y., Panagiotou, G., Mougios, V., Brinkoetter, M., Vamvini, M. T., Schneider, B. E. & Mantzoros, C. S. (2012). FNDC5 and irisin in humans: I. Predictors of circulating concentrations in serum and plasma and II. mRNA expression and circulating concentrations in response to weight loss and exercise. Metabolism, 61, 1725-1738.
Hyatt, J. P., Nguyen, L., Hal,l A. E., Huber, A. M., Kocan, J. C., Mattison, J. A., de Cabo, R., LaRocque, J. R., & Talmadge, R. J. (2016). Muscle-Specific Myosin Heavy Chain Shifts in Response to a Long-Term High-fat/High Sugar Diet and Resveratrol Treatment in Nonhuman Primates. Frontiers in Physiology, 2, 7:77.
Kurdiova, T., Balaz, M., Vician, M., Maderova, D., Vlcek, M., Valkovic, L., Srbecky, M., Imrich, R., Kyselovicova, O., Belan, V., Jelok, I., Wolfrum, C., Klimes, I., Krssak, M., Zemkova, E., Gasperikova, D., Ukropec, J., & Ukropcova, B. (2014). Effects of obesity, diabetes and exercise on Fndc5 gene expression and irisin release in human skeletal muscle and adipose tissue: in vivo and in vitro studies. The Journal of Physiology, 592(5), 1091-107.
Lee, P., Linderman, J. D., Smith, S., Brychta, R. J., Wang, J., Idelson, C., Perron, R. M., Werner, C. D., Phan, G. Q., Kammula, U. S., Kebebew, E., Pacak, K., Chen, K. Y., & Celi, F. S. (2014). Irisin and FGF21 are cold induced endocrine activators of brown fat function in humans. Cell Metabolism, 19, 302–309.
Moreno-Navarrete, J. M., Ortega, F., Serrano, M., Guerra, E., Pardo, G., Tinahones, ., Ricart, W., & Fernández-Real, J. M. (2013). Irisin is expressed and produced by human muscle and adipose tissue in association with obesity and insulin resistance. The Journal of clinical endocrinology and metabolism, 98(4), E769-78.
Norheim, F., Langleite T. M., Hjorth, M., Holen, T., Kielland, A., Stadheim, H. K., Gulseth, H. L., Birkeland, K. I., Jensen, J., & Drevon, C. A. (2014). The effects of acute and chronic exercise on PGC-1α, irisin and browning of subcutaneous adipose tissue in humans. http://onlinelibrary.wiley.com/journal/10.1111/(ISSN)1742-4658, 281(3), 739-49.
Widrick, J. J., Norenberg, K. M., Romatowski, J. G., Blaser, C. A., Karhanek, M., Sherwood, J., Trappe, S. W., Trappe, T. A., Costill, D. L., & Fitts, R. H. (1998). Force-velocity-power and force-pCa relationships of human soleus fibers after 17 days of bed rest. Journal of Applied Physiology, 85(5), 1949-1956.
Yu, J., Zheng, J., Liu, X. F., Feng, Z. L., Zhang, X. P., Cao, L. L., Zhou, Z. P. (2016). Exercise improved lipid metabolism and insulin sensitivity in rats fed a high-fat diet by regulating glucose transporter 4 (GLUT4) and musclin expression. Brazilian Journal of Medical and Biological Research, 49(5), e5129.